


Scientific journal
Modern high technologies
ISSN 1812-7320
"Перечень" ВАК
ИФ РИНЦ = 1,279
ORIGIN OF LYMPHATIC ENDOTHELIUM. I. HUMAN EMBRIOGENESIS

Известно множество гипотез возникновения лимфатического русла в эмбриогенезе человека и животных. Их сводят к двум диаметрально противоположным точкам зрения: 1) согласно F. Sabin [9], выросты передних кардинальных вен отделяются и сливаются в яремные лимфатические мешки (ЛМ), а из них вырастают грудные протоки, лимфатический эндотелий развивается из венозного эндотелия [8]. По мнению S. Putte [7], первичные венозные сети превращаются в ЛМ, которые изначально представлены лимфатическими сплетениями; 2) по утверждению O. Kampmeier [5], часть эмбриональных вен исчезает (куда ?), на их месте появляются мезенхимные щели (откуда ?), они сливаются в первичные лимфатические коллекторы, их эндотелий образуется из мезенхимных клеток. Представления F.Lewis [6] занимают промежуточное положение: лимфатические сосуды формируются благодаря более длительной серии изоляций, чем просто ответвления вен F.Sabin, частью из вен, частью из лимфатических щелей с эндотелиальной выстилкой, которые выпячиваются из вен, а затем сливаются. Но F. Lewis не объяснил механику образования лимфатических щелей.
Результаты современных исследований с применением стволовых клеток и методов экспрессии генов также трактуются двояко, с диаметрально противоположными выводами о венозном или мезенхимном происхождении лимфатического эндотелия в эмбриогенезе животных [3, 4, 10-13].
По моим данным [1-2], в процессе расширения эмбриональные вены с эндотелиальными стенками охватывают артерии с более толстыми стенками. Иначе говоря, артерии с адвентициальной оболочкой инвагинируют в полость вен вместе с участками их эндотелиальных стенок. В результате периферическая часть первичных вен разделяется на карманы. Расширяясь, они отшнуровываются от центрального канала матричной вены (сужение и перерыв их сообщений), отделяются в виде лимфатических щелей вместе с эндотелиальной выстилкой предшествующих венозных карманов. Эндотелий вторичной вены (центральный канал первичной вены с магистральным кровотоком) утолщается и окружается тонкой адвентициальной оболочкой. Лимфатические щели расширяются, перегородки между ними истончаются и щели сливаются в ЛМ. Притоки первичных вен, которые впадали в карманы венозных коллекторов, также выключаются из кровотока и становятся лимфатическими сосудами с таким же тонкими эндотелиальными стенками, как в прешествующих им венах. В их числе – грудные протоки, поясничные и другие стволы. В морфогенезе ЛМ, прежде всего яремных, мной обнаружен несколько иной механизм выключения вен из кровотока: мелкие артерии сдавливают участки мелких притоков крупных вен, промежуточные участки мелких вен расширяются и обосабливаются в виде лимфатических щелей. Мезенхима на стадии закладки ЛМ отсутствует в их окружении, она уже преобразовалась в соединительную ткань с очень рыхлой сетью тонких ретикулярных волокон, которая окружает кровеносные микрососуды.
Цель исследования: показать однотипность и преемственность эндотелиальной выстилки венозных карманов и последующих лимфатических щелей на примере морфогенеза забрюшинного ЛМ у эмбрионов человека.
Материалы и методы исследования
20 эмбрионов человека 5-30 мм теменно-копчиковой длины (4-8 нед) были фиксированы в жидкости Буэна и залиты в парафин. Их серийные срезы толщиной 5-7 мкм в 3 основных плоскостях были окрашены гематоксилином и эозином, смесью Маллори, альциановым синим (рН = 2,0-3,0) в комбинации с ШИК-реакцией, толуидиновым синим (рН = 1,0-5,0), а также импрегнированы нитратом серебра по Карупу на ретикулярные волокна соединительной ткани.
Результаты исследования и их обсуждение
У эмбрионов человека 14-15 мм длины (начало 7-й нед) верхний интерсубкардинальный анастомоз соединяется с мезокардинальными венами и резко расширяется с образованием субкардинального венозного синуса между почками и надпочечниками. Тонкие эндотелиальные стенки синуса при этом сильно деформируются. Их участки с прилегающими артериями мезонефроса, обладающими более толстыми стенками с адвентициальной оболочкой, и межсосудистой соединительной тканью инвагинируют в просвет синуса (рис. 1). Инвагинации разных размеров расчленяют периферическую часть субкардинального синуса, прежде всего в дорсальном отделе, на полиморфные венозные карманы. У эмбрионов 17-20 мм длины (6,5-7 нед) субкардинальный венозный синус разделяется на две части: 1) центральная, с магистральным кровотоком и очень тонкой адвентициальной оболочкой – ствол левой почечной вены и почечная часть нижней полой вены; 2) периферическая, в виде скопления полиморфных лимфатических щелей с очень тонкой эндотелиальной выстилкой – закладка забрюшинного ЛМ (рис. 2). В ее каудальной части преобладают лимфатические щели, которые формируются из мелких вен субкардинального сплетения. У эмбрионов 7-8 нед межщелевые перегородки истончаются и разрываются (рис. 3). В результате образуется и постепенно расширяется крупная полость забрюшинного ЛМ. Он окружает ствол левой почечной вены и прилегающую часть нижней полой вены, имеет очень тонкие эндотелиальные стенки. Дольше сохраняются те более толстые межщелевые перегородки, покрытые тонким эндотелием, в которых сохраняются артерии мезонефроса. На данном уровне первичные почки дегенерируют, с ними – и большинство их артерий, кроме тех, которые преобразуются в артерии дефинитивных органов, например почек, или их закладок, например, артерии тел Цукеркандля.
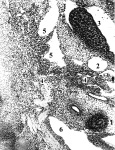
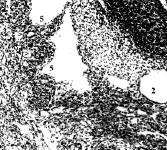
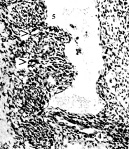
Рис. 1. Эмбрион человека 14 мм длины (начало 7-й недели), сагиттальные срезы: 1 – правая почка; 2 – мезонефрос; 3 – гонада; 4 – артерия мезонефроса; 5 – венозные карманы, инвагинации и перегородка в полости субкардинального венозного синуса; 6 – правая нижняя мезокардинальная вена. Стрелками показаны ветви артерий мезонефроса, которые инвагинируют в полость синуса. Хорошо видна эндотелиальная выстилка стенок синуса и инвагинаций его дорсальной стенки. Гематоксилин и эозин. Ув.: А – 120; Б – 200; В – 300
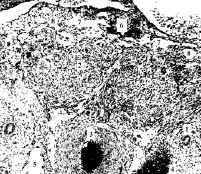
Рис. 2. Эмбрион человека 20 мм длины (7 недель), поперечный срез: 1 – мочеточник; 2 – тела Цукеркандля; 3 – брюшная аорта; 4 – нижняя полая вена; 5 – левый поясничный ствол; 6 – ствол левой почечной вены; 7,8 – забрюшинный лимфатический мешок и лимфатические щели его закладки, мешок и щели имеют эндотелиальную выстилку. Гематоксилин и эозин. Ув. 100
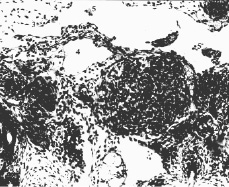
Рис. 3. Эмбрион человека 25 мм длины (7,5 недель), поперечный срез: 1 – почка; 2 – тела Цукеркандля; 3,4,5 – забрюшинный лимфатический мешок, лимфатические щели и остатки межщелевых перегородок его закладки; 6 – микрососуд в сохранившейся межщелевой перегородке. Хорошо видна эндотелиальная выстилка стенок мешка и щелей. Гематоксилин и эозин. Ув. 200
Заключение
Эндотелиальная выстилка субкардинального венозного синуса и его карманов, образующихся из них лимфатических щелей и забрюшинного ЛМ определяется у эмбрионов человека постоянно, имеет одинаковые толщину и строение, эндотелий левой почечной вены – большую толщину при более частом размещении клеточных ядер. На этом основании я утверждаю: лимфатический эндотелий возникает из эндотелия той части первичного венозного русла, которая выключается из кровотока у эмбрионов человека второго месяца путем образования венозных карманов и их обособления в виде лимфатических щелей. Щели затем сливаются в ЛМ. Дифференциация лимфатического и венозного эндотелиев, судя по их толщине и строению, происходит по градиенту кровяного давления. Эти параметры еще больше у эндотелия артерий, как и артериальное давление.
Библиографическая ссылка
Петренко В.М. ПРОИСХОЖДЕНИЕ ЛИМФАТИЧЕСКОГО ЭНДОТЕЛИЯ. I. ЭМБРИОГЕНЕЗ ЧЕЛОВЕКА // Современные наукоемкие технологии. 2014. № 1. С. 71-74;URL: https://top-technologies.ru/en/article/view?id=33634 (дата обращения: 18.07.2026).