


Scientific journal
Modern high technologies
ISSN 1812-7320
"Перечень" ВАК
ИФ РИНЦ = 1,279
Ability of histidine, triptoрhan, tyrosine, trimetazidin, mildronat and blood serum to reduce -adrenoblokatory effect of ozone

Методика исследований
Опыты проведены на 197 продольных полосах (длиной 6-8 мм и шириной 2-3 мм) рога матки 28 крыс (фаза метаэструса или диэструса). Регистрацию сократительной активности полосок и оценку их b-адренореактивности проводили по [11] на «Миоцитографе» при 38оС, пассивной аэрации раствора Кребса, при перфузии 0,7 мл/мин и исходной нагрузке 4,9 мН. Выполнено 12 серий: в сериях 1 и 2 использовали спонтанно активные полоски, а в сериях 3-12 - полоски с вызванной гиперкалиевым (60 мМ KCl) раствором Кребса тонической активностью. Стандартный раствор Кребса (рН-7,4), содержал (в мМ) NaCl-136, КCl-4,7, CaCl2-2,52, MgCl2-1,2, KH2PO4-0,6, NaHCO3-4,7, C6H12O6-11. На его основе готовили озонированный раствор Кребса. Для этого стандартный раствор Кребса в течение 20 минут барботировали озоно-кислородной смесью (ее получали на озонаторе типа «Озон-М-50») при концентрации озона 5х10-3 г/л. По расчетам [2], концентрация озона в таком растворе на протяжении 1-3 часа удерживается на уровне 5х10-7 г/мл. Этот раствор разводили стандартным раствором Кребса в 10 раз; в нем концентрация озона достигала 5х10-8 г/мл, т.е. близкой к терапевтической [2,3,16]. В работе использовали триметазидин (Франция), милдронат (Латвия), гистидин и триптофан (Венгрия), тирозин и адреналина гидрохлорид (Россия). В качестве источника эндогенного сенсибилизатора β-адренорецепторов применяли 100-кратно разведенную (стандартным раствором Кребса) сыворотку венозной крови 6 небеременных женщин (фолликулярная фаза). Различия между показателями оценивали по критерию Стьюдента, считая их достоверными при р<0,05 .
Результаты исследования
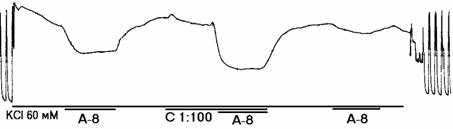
В сериях 1 и 2 в опытах со спонтанно активными полосками изучали влияние гистидина (3х10-6 г/мл) и триметазидина (10-6 г/мл) на b-адреноблокирующий эффект озонированного раствора Кребса (далее - озон), при его концентрации 5х10-8 г/мл. Для этого (рис. 1) проводили 4-кратное (по 10 мин.) тестирование адреналином (10-8 г/мл): до озона, на фоне озона, на фоне озона и гистидина (триметазидина) и после их удаления. При этом озон воздействовал 30 минут: с 11-й по 20-ю минуту - совместно с адреналином, а с 21-й по 30-ю минуту - совместно с адреналином и исследуемым веществом. Установлено (рис. 1, панели а, б), что озон не влиял на сократимость полосок, но достоверно снижал их β-адренореактивность, а гистидин и триметазидин восстанавливали ее. Так, в серии 1 суммарная сократительная активность при 1-м тестировании адреналином составила 76,9±10,4%* от исходного уровня (*- изменения носят достоверный характер, р<0,05), на фоне озона - 110±14,8%, при 2-м тестировании адреналином (т.е. адреналин + озон) - 85,9±13,2%, при 3-м (адреналин + озон + гистидин) - 61,5±8,47%*, а при 4-м (адреналин) - 66,7±9,8%*; близкие значения получены в серии 2, т.е. с триметазидином - соответственно 73,1±11,4%*, 122±16,8%, 94,4±13,9%, 81,1±9,20% и 79,1±10,4%.
В сериях 3-6 исследовали влияние гистидина (3х10-6 г/мл), триметазидина (10-6 г/мл), сыворотки (1:100) крови и озона (5х10-8 г/мл) на b-адренореактивность полосок матки в условиях тонической активности, вызванной гиперкалиевым (60 мМ KCl) раствором Кребса. На ее фоне проводили три (по 10 минут) тестирования адреналином (10-8 г/мл): до, во время и после воздействия исследуемого фактора. Установлено (табл.1, рис.2, а-е) что адреналин при 1-м тестировании достоверно снижал калиевую контрактуру, гистидин незначительно снижал ее (рис.2, а), а триметазидин, сыворотка крови и озон не изменяли ее. При 2-м тестировании, т.е. на фоне гистидина (рис.2, а), триметазидина (рис.2 б) и сыворотки крови (рис.2,в) релаксирующий эффект адреналина достоверно увеличился, а на фоне озона, наоборот, уменьшился (рис.2, г, д, е). При 3-м тестировании релаксирующий эффект адреналина восстанавливался до исходного уровня. Следовательно, β-адренореактивность деполяризованных миоцитов матки крысы под влиянием гистидина, триметазидина и эндогенного сенсибилизатора β-адренорецепторов существенно возрастает, а под влиянием озона снижается.
Таблица 1. Влияние различных факторов на β-адренореактивность продольных полосок рога матки небеременных крыс, деполяризованных гиперкалиевым (60 мМ КСl) раствором Кребса, в % к фоновому уровню.
|
Фактор |
Этапы эксперимента |
|||
|
2 |
4 |
5 |
7 |
|
|
KCl+адреналин, 10-8 г/мл |
KCl+исследуемый фактор |
KCl+исследуемый фактор+адреналин |
KCl+адреналин 10-8 г/мл |
|
|
Гистидин, (3х10-6 г/мл, n=12) |
88,0±6,61 |
84,0±4,89* |
40,0±8,56*а |
92,0±4,58 с |
|
Триметазидин, (10-6 г/мл, n=9) |
62,1±4,57* |
92,1±3,34 |
32,8±5,4*а |
77±8,1* с |
|
Сыворотка крови (1:100, n=10) |
91±5,6 |
89±7,6 |
45±6,2 *а |
95±8,3 с |
|
Озонированный раствор Кребса, (5х10-8 г/мл, n=54) |
66,6±8,21* |
105±7,54 |
101±9,81а |
67,4±4,63* с |
Примечание: * - различия с исходным уровнем достоверны, (р<0,05); а, с, - означают достоверность различий (р<0,05) между значениями 2-го и 5-го (а), 5-го и 7-го (с).
Таблица.2. Влияние различных сенсибилизаторов b-адренорецепторов на b-адренореактивность продольных полосок рога матки небеременных крыс, сниженную озонированным раствором Кребса 5х10-8 г/мл в условиях калиевой (60 мМ КСl) контрактуры, в % к фоновому уровню.
|
Сенсибилизатор |
Этапы эксперимента |
||||
|
2 |
4 |
5 |
6 |
8 |
|
|
KCl +адреналин, 10-8 г/мл |
KCl+ОРК |
KCl+ОРК +адреналин |
KCl+ОРК +адреналин +сенсибилизатор |
KCl+адреналин, 10-8 г/мл |
|
|
Гистидин (3х10-6 г/мл, n=9). |
76,8±5,34* |
103±4,91 |
103±7,86d |
33,3±5,42*аe |
54,7±5,73* bс |
|
Триптофан (10-6 г/мл, n=9) |
47,4±7,23* |
111±5,84 |
108±7,03d |
45,4±8,01*e |
72,8±6,95*с |
|
Тирозин (2х10-6 г/мл, n=9) |
62,0±8,50* |
107±1,65 |
101±6,04d |
44,0±3,76*e |
80,3±5,55с |
|
Триметазидин (10-6 г/мл,n=9) |
72,7±2,78* |
108±1,79 |
109±2,31d |
77,9±6,82*e |
81,6±2,39 |
|
Милдронат, (10-5 г/мл, n=9). |
71,0±1,36* |
114±3,76 |
107±2,43d |
82,0±2,19e |
73,0±0,97* |
|
Сыворотка крови (1:100, n=9) |
60,1±4,44* |
108±3,94 |
101±7,62d |
26,4±4,37*аe |
67,6±4,83*с |
Примечание: * - различия с исходной тонической активностью (т.е. с 1-ым этапом эксперимента) достоверны, р<0,05; а, b, с, d, e - различия между значениями соответственно 2-го и 6-го (а), 2-го и 8-го (b), 6-го и 8-го (с), 2-го и 5-го (d), 5-го и 6-го (e) этапами эксперимента достоверны, р<0,05.
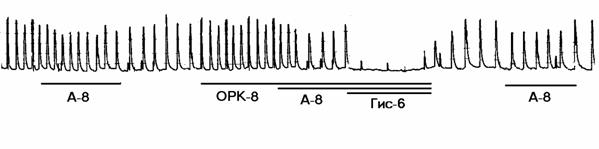
а
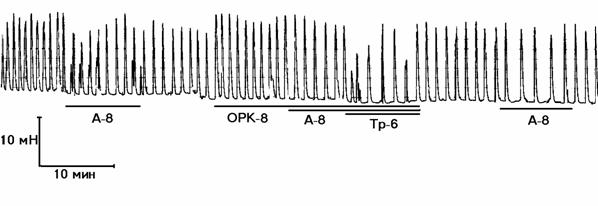
б
Рисунок 1. Механограммы продольных полосок рога матки небеременной крысы, демонстрирующие b-адреноблокирующее влияние озонированного раствора Кребса (5х10-8 г/мл) и снижение этого влияния с помощью гистидина (панель а) и триметазидина (панель б). Горизонтальные линии под механограммами показывают момент воздействия озонированного раствора Кребса (5х10-8 г/мл, ОРК-8), триметазидина (10-6 г/мл, Тр-6), гистидина (3х10-6 г/мл; Гис-6) и адреналина (10-8 г/мл, А-8). Калибровка 10 мН, 10 мин.
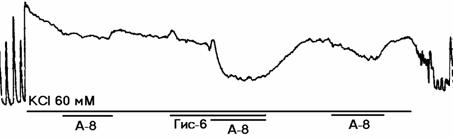 а
а
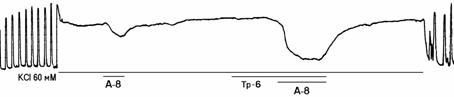 б
б
 в
в
 г
г
 д
д
 е
е
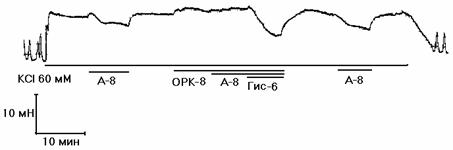
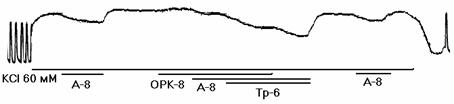
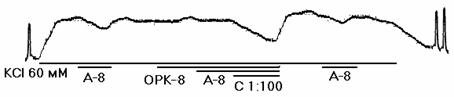
Рисунок 2. Механограммы продольных полосок рога матки небеременной крысы, демонстрирующие на фоне калиевой контрактуры отсутствие утероингибирующего и наличие b-адреносенсибилизирующего эффектов гистидина (панель а), триметазидина (панель б), сыворотки крови небеременных женщин (панель в), а также b-адреноблокирующее влияние озонированного раствора Кребса (панели г, д, е) и его снижение гистидином (панель г), триметазидином (панель д) и сывороткой крови небеременных женщин (панель е). Горизонтальные линии под механограммами показывают момент воздействия гиперкалиевого раствора Кребса (60 мМ КСl), озонированного раствора Кребса (5х10-8 г/мл, ОРК-8), гистидина (3х10-6 г/мл; Гис-6), триметазидина (10-6 г/мл, Тр-6), сыворотки крови (С1:100) и адреналина (10-8 г/мл, А-8). Калибровка 10 мН, 10 мин.
В сериях 7-12 на продольных полосках матки крысы изучали влияние гистидина (3х10-6г/мл), триптофана (10-6 г/мл), тирозина (2х10-6 г/мл), триметазидина (10-6 г/мл), милдроната (10-5 г/мл) и сыворотки (1:100) крови на b-адреноблокирующий эффект озона (5х10-8 г/мл). Для этого на фоне калиевой контрактуры проводили 4 (по 10 минут) тестирования адреналином (10-8 г/мл): до озона, на его фоне, на фоне озона и исследуемого фактора и после их удаления. Установлено (табл.2, рис 2, г-е), что адреналин при 1-м тестировании достоверно снижал калиевую контрактуру, а озон не изменял ее, но существенно снижал релаксирующий эффект адреналина (2-тестирование). Гистидин (рис. 2, г), триптофан, тирозин, триметазидин (рис. 2, д), милдронат и сыворотки крови (рис 2, е) существенно и обратимо повышали релаксирующий эффект адреналина.
Следовательно, гистидин, триптофан, тирозин, триметазидин, милдронат и эндогенный сенсибилизатор β-адренорецепторов независимо от уровня мембранного потенциала миоцитов уменьшают проявление β-адреноблокирующего действия озона.
Обсуждение результатов
С учетом механизма β-адренорецепции [6,15,17,18], полагаем, что озон снижает β-адренореактивность миоцитов за счет уменьшения сродства β-адренорецепторов к агонисту (в результате активации фосфорилирования этих рецепторов?) или разобщения передачи сигнала внутрь клетки. Не исключено, что такое действие озона может быть использовано при лечении артериальной гипертензии (например, в комплексе с β-адреноблокаторами). С другой стороны, результаты исследования показывают, что озон может вызывать нежелательные побочные эффекты, оказывая b-адреноблокирующий эффект. Это может быть противопоказанием для озонотерапии или выполнения озоноопасных работ. В этих случаях для уменьшения негативного действия озона, как показали результаты исследования, вероятно, целесообразно использовать β-адреносенсибилизаторы прямого действия, т.е. гистидин, триптофан, тирозин, предуктал и милдронат, эффект которых, возможно, обусловлен восстановлением сродства β-адренорецепторов к агонистам (вследствие активации фосфатазы, осуществляющей дефосфорилирование β-адренорецепторов?) или передачи сигнала от β-адренорецепторов внутрь клетки. Как известно [10,13], содержание эндогенного сенсибилизатора β-адренорецепторов в крови зависит от пола (у женщин выше, чем у мужчин) и этапа репродуктивного процесса (у беременных женщин выше, чем у небеременных). Наши данные о способности сыворотки крови восстанавливать сниженную под влиянием озона β-адренореактивность позволяет предположить, что реакция организма на воздействие озона будет зависеть от содержания в крови эндогенного сенсибилизатора β-адренорецепторов - чем оно выше, тем меньше должен проявляться β-адреноблокирующий эффект озона. Таким образом, при озонотерапии и проведении озоноопасных работ следует учитывать содержание в крови эндогенного сенсибилизатора b-адренорецепторов и его компонентов (гистидина, триптофана и тирозина). В целом, обнаружение принципиально нового свойства у гистидина, тирозина, триптофана, триметазидина, милдроната и эндогенного сенсибилизатора β-адренорецепторов расширяет перспективу разработки новых методов лечения патологии внутренних органов, обусловленной недостаточностью b-адренергических влияний. С другой стороны, способность сыворотки крови, гистидина, триптофана и тирозина уменьшать β-адреноблокирующий эффект озона позволяют говорить об участии эндогенного сенсибилизатора β-адренорецепторов и его компонентов в поддержании оптимальной β-адренореактивности клеток организма в различных условиях обитания человека и животных, в том числе в условиях повышенного содержания озона в окружающей среде. С этих позиций дальнейшее изучение физиологической роли эндогенного сенсибилизатора β-адренорецепторов и его компонентов (гистидина, триптофана и тирозина) следует рассматривать как одно из перспективных направлений в физиологии и медицины.
Список литературы
- Белянин И.И., Шмелев Е.И. //Проблемы туберкулеза.1998. № 1. С. 30.
- Бояринов Г.А., Соколов В.В. Озонированное искусственное кровообращение. Н. Новгород: «Покровка», 1999. 317 с.
- Густов А.В., Котов С.А., Конторщикова К.Н., Потехина Ю.П. Озонотерапия в неврологии. Н. Новгород: Литера, 1999. 178с.
- Качалина Т.С., Гречканев Г.О., Шахова Н.М. //Озон и методы эфферентной терапии в медицине: Тез. докл. конф. Н. Новгород: НГМА, 1998. С. 50.
- Ноздрачев А.Д., Туманова Т.В., Дворянский С.А. и др. // Доклады РАН. 1998. Т. 363, № 1. С. 133.
- Сергеев П.В., Шимановский Н.Л., Петров В.И. Рецепторы физиологически активных веществ. Волгоград: «Семь ветров», 1999. 640с.
- Сизова Е.Н., Циркин В.И., Дворянский С.А. // Росс. физиол. журн. им. И.М. Сеченова. 2002.Т. 88, № 7. С. 856.
- Сизова Е.Н., Циркин В.И., Костяев А.А. и др. //Биология - наука XXI века: Сб. тезисов 6-й Пущинской школы-конференции молодых ученых. Пущино, 2002. С. 139.
- Сизова Е.Н., Циркин В.И., Подтетенев А.Д. и др. // Росс. кардиол. журн. 2002. № 2 (34). С. 50.
- Циркин В.И., Дворянский С.А. Сократительная деятельность матки (механизмы регуляции). - Киров: КГМА, 1997. 270 с.
- Циркин В.И., Дворянский С.А., Ноздрачев А.Д. и др. // Доклады РАН.-1996. Т. 351, № 4. С. 565.
- Циркин В.И., Дворянский С.А., Джергения С.Л. и др. // Физиология человека. 1997. Т. 23, № 5. С 85.
- Циркин В.И., Дворянский С.А., Ноздрачев и др. //Доклады РАН. - 1997. Т. 352, № 1. С. 124.
- Циркин В.И., Сизова Е.Н., Подтетенев А.Д. и др. //Росс. кардиол. журн., 2002.№ 1 (33). С. 45.
- Циркин В.И., Трухина С.И. Физиологические основы психической деятельности и поведения человека.- М.: Медицинская книга, 2001. 524 с.
- Яворская В.А., Малахов В.А., Хвисюк В.В. и др. //Мiсцеве та парентеральне використання озонотерапii в медицинi: Сб. наук. робiт.-Харькiв, 2001. С. 45.
- Chuang Tsu Tshen, Sallese M, Ambrosini G. et al. // J. Biol. Chemistry. 1992.V.267, № 10. Р. 6886.
- Haussdorff W., Caron M., Lefkowitz R. // FASEB Journal, 1990. V. 4, № 11. Р. 2881
Библиографическая ссылка
Сизова Е.Н., Циркин В.И., Туманова Т.В., Костяев А.А. СПОСОБНОСТЬ ГИСТИДИНА, ТРИПТОФАНА, ТИРОЗИНА, ТРИМЕТАЗИДИНА, МИЛДРОНАТА И СЫВОРОТКИ КРОВИ УМЕНЬШАТЬ b–АДРЕНОБЛОКИРУЮЩИЙ ЭФФЕКТ ОЗОНА // Современные наукоемкие технологии. 2004. № 3. С. 21-260;URL: https://top-technologies.ru/en/article/view?id=21746 (дата обращения: 03.07.2026).