



Объёмы переработки природных биологических (растительных и животных) тканей дроблением, измельчением, резанием с трудом поддаются оценке. Эти процессы являются наиболее энергоёмкими в сельхозпереработке, пищевой, лесоперерабатывающей промышленностях и т.д. Если на процессы дезинтеграции, по некоторым оценкам, человечество затрачивает до 8 % всей вырабатываемой энергии, то не менее трети её приходится на переработку сырья биологического происхождения. Растительное сырье относится к категории трудноизмельчаемых материалов благодаря прочной волокнистой структуре, содержащей клетчатку в различных её формах.
Наряду с тканями и клетками, играющими в теле растения (как сооружения) роль, аналогичную роли заполнения в железобетонных конструкциях, в растениях имеются ткани и клетки, соответствующие арматуре железобетона: стержням, проволокам, пронизывающим (а иногда и обматывающим) основную массу. Ткани и клетки-одиночки подобного рода можно назвать арматурными, или механическими, а совокупность их – арматурой, стереомом, или механической системой. Арматура растений может быть представлена, нередко уже в пределах одной особи, тремя различными типами тканей и клеток [1]: склеренхимой, колленхимой и каменистыми клетками (склереидами).
Основными чертами строения, характерными для всех типов арматуры, являются: 1) мощное утолщение клеточных стенок, 2) тесное, за редкими исключениями, смыкание между клетками, 3) отсутствие перфораций в клеточных стенках. Живое содержимое клеток рано отмирает (в типичных склероидах и в склеренхиме), реже сохраняется долго (в клетках колленхимы).
Сильно утолщенные клеточные стенки арматурных тканей обладают высокими физико-механическими качествами. Располагается арматура в органах растений согласно принципам обеспечения механической прочности, в широком смысле слова, с наивозможно малой затратой материала.
Арматурной тканью, наиболее важной и по распространенности у растений, и по относительной массе материала клеточных стенок, и по значению для прочности растений, является склеренхима, в типичной ее форме – ткань, состоящая из клеток, плотно сомкнутых, прозенхимных (т.е. удлиненных и заостренных на двух противоположных концах), толстостенных, с немногочисленными узкими простыми щелевидными порами, расположенными длинной осью под острым углом к продольной оси клетки. Клетки склеренхимы могут быть названы толстостенными волокнами. В технике их называют «волоконцами», или «элементарными волокнами». При полном своем развитии клетки склеренхимы обычно не имеют живого содержимого, и полости их заполняются воздухом. К этому времени клеточные стенки в большинстве случаев одревесневают. Сравнительно редко они остаются неодревесневшими.
Материал клеточных стенок типичной склеренхимы по прочности на разрыв (σв) и по величине предела упругости (σупр) близок к строительной стали. В отличие от строительной стали, удлинения склеренхимы упруги почти до разрыва, притом они значительно более высоки, нежели у стали. В связи с этим у склеренхимы величины модуля упругости (Е) гораздо ниже, а величины работы деформации («живого» упругого сопротивления) на единицу объема (ауп) выше, нежели у стали. По способности противостоять без остающихся деформаций динамическим нагрузкам склеренхима значительно превосходит строительную сталь.
Решающую роль в процессе разрушения растительного сырья играют физико-химические факторы [1]: поверхностные явления на границе раздела фаз, контактные взаимодействия между ними и, как следствие, непрерывное образование и разрушение трехмерных структур, агрегатов из частиц непосредственно в ходе процессов получения однородных высококонцентрированных дисперсных систем [5, 6, 7].
Целью исследований являлось выявление закономерностей изменения прочностных свойств тканей биологического происхождения, подвергнутых физическим воздействиям (в первую очередь – тепловым), а также обезвоживанию.
В качестве объекта исследования был выбран корень лопуха обыкновенного как перспективный вид сырья широкого диапазона использования.
Для проведения экспериментов было подготовлено 100 образцов корня лопуха большого (Arctium. lappa) кубической формы с длиной ребра 10 мм. Все подготовленные образцы подвергались конвективно-радиационной сушке при температуре до 60 °С. Кривая сушки образцов корня лопуха обыкновенного представлена на рис. 1.
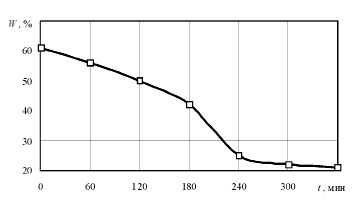
Рис. 1. Кривая сушки образцов корня лопуха обыкновенного
Для определения прочностных параметров при сжатии и сдвиге партии образцов различной влажности испытывались на приборе «Структурометр СТ–1». Испытания проводились при квазистатическом нагружении сжатием и сдвигом с различными скоростями. Образцы корня лопуха подвергались деформации и разрушению как вдоль волокон, так и поперек при различных скоростях перемещения столика V=0,33∙10-3 м/с, V=0,83∙10-3 м/с, V=1,17∙10-3 м/с и V=1,67∙10-3 м/с; разрушение образца производили до достижения заданного значения предельного усилия нагружения F=60H. Это значение было установлено при проведении пробных экспериментов, оно является достаточным для разрушения любого образца данного типа.
В процессе сжатия и сдвига исследуемых образцов в устройстве при каждом шаге измерений регистрировались следующие экспериментальные данные: N – число измерений (количество перемещений столика); F – усилие нагружения, Н; t – время, с; h – деформация образца (значение перемещения столика), мм. При помощи установленной на персональном компьютере программы полученные данные считывались с прибора и записывались в текстовые файлы для просмотра. Информация была представлена в виде таблиц усилий. Содержимое таблиц иллюстрируется графиками F=f(N).
Были получены зависимости предельных напряжений разрушения и сдвига от влажности образцов. Предельное напряжение разрушения σпр, МПа, и предельное напряжение сдвига τпр, МПа, вычислялись по формуле
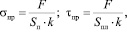
где F – экспериментальное усилие, Н; Sп – площадь поперечного сечения образца, нормального направлению нагрузке, м2; Sпл – площадь сечения образца в направлении сдвиговой нагрузки, м2; k – коэффициент усушки.
На рис. 2 – 5 представлены зависимости предельных напряжений разрушения при сжатии и сдвиге образцов корня лопуха при их различной влажности.
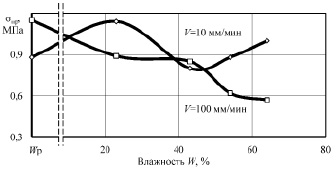
Рис. 2. Зависимость предельных напряжений разрушения sпр от влажности W образцов корня лопуха при сжатии поперек оси
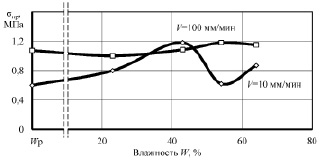
Рис. 3. Зависимость предельных напряжений разрушения sпр от влажности W образцов корня лопуха при сжатии вдоль оси
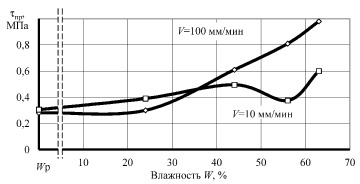
Рис. 4. Зависимость предельных напряжений сдвига tпр от влажности W образцов корня лопуха при сдвиге вдоль оси
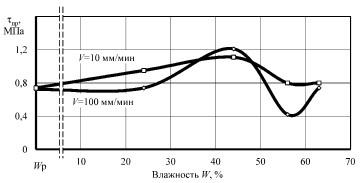
Рис. 5. Зависимость предельных напряжений сдвига tпр от влажности W образцов корня лопуха при сдвиге поперек оси
Анализ графиков показывает наличие характерных точек и участков. Исходная (нативная) влажность образцов составила 62 %. Понижение влажности до 56 % привело к минимальной прочности при сжатии и сдвиге вдоль волокон и повышению прочности до максимальной при сжатии вдоль оси корня при скорости нагружения 100 мм/мин. Очевидно, что такое деформационное поведение растительной ткани связано с исчезновением тургора, внутреннего давления в клетках, что приводит к снижению упругости ткани, а следовательно, понижению внутренней энергии вещества. Увеличение прочности при такой влажности на повышенной скорости деформирования, видимо, связано с исчезновением релаксационных процессов, когда напряжения не успевают более равномерно распространиться в объёме образца.
Влажность материала, близкую к 44 %, можно назвать первой критической точкой. При этой влажности происходит резкое изменение скорости сушки, практически заканчивается уменьшение размеров образцов (усушка), а на кривых прочности наблюдаются экстремумы, либо же возрастающая прочность. Несомненно, что при этой влажности заканчивается удаление физически связанной влаги из образцов, преимущественно из межклеточных пространств [8].
При квазистатическом сжатии вдоль оси, а также при сдвиге как вдоль, так и поперек волокон при влажности 44 % прочность образцов достигает максимума. Удаление влаги приводит к формированию более прочного адгезионного взаимодействия между клеточными структурами растительной ткани, несмотря на отсутствие активной воды, внутренняя энергия вещества повысилась, связанная физико-химически с макромолекулами растительной ткани вода повышает не только межклеточную, но и межструктурную прочность образцов.
Дальнейшее обезвоживание образцов приводит к стабильному снижению прочности до минимальной при равновесной влажности, и только при сжатии поперек волокон наблюдается рост прочности при повышенной скорости приложения нагрузки. Природа прочности растительной ткани при минимальной влажности, скорее всего, перестает носить адгезионный характер и определяется прочностью целлюлозного каркаса при растяжении, в частности – поперечными связями между макромолекулами биополимеров.
Следует отдельно рассмотреть влияние скорости деформирования образцов на их прочность. В нашем случае испытания проведены при скоростях 10 и 100 мм/мин. При нативной влажности, когда образцы сжимались вдоль оси и при сдвиге поперек оси, десятикратное увеличение скорости привело к более чем двукратному повышению предела прочности. При влажности 56 % разность прочностей при различных скоростях увеличилась независимо от вида испытаний. После удаления активной воды разница в деформационном поведении образцов значительно снизилась, особенно при сдвиге.
Интересным представляется сравнение динамики изменения прочностных свойств растительной и животной ткани, например – костной. Структурной и функциональной единицей костной ткани является остеон. Остеон – это система концентрических цилиндрических костных пластинок, вставленных друг в друга. Между пластинками остеона находятся костные клетки. Остеоны ориентированы по направлению действия наибольших нагрузок, между ними располагаются вставочные костные пластинки. По данным А.А. Соколова (МТИММП, кафедра технологического оборудования мясокомбинатов), на прочностные свойства кости значительно влияет термическое воздействие. График зависимости ударной вязкости от продолжительности термической обработки представлен на рис. 6.
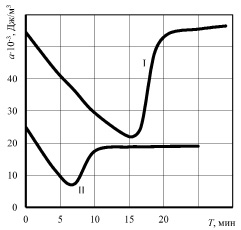
Рис. 6. Зависимость ударной вязкости костной ткани от продолжительности термической обработки (по результатам А.А. Соколова): I – говяжья реберная кость, II – свиная реберная кость
Как объяснить резкое снижение прочности кости в результате термической обработки? Если подходить к этому вопросу с позиции адгезионной природы прочности биологических тканей [2, 3, 4], то картина становится достаточно ясной. Белки оссеин и коллаген выступают в костной ткани связующим межклеточным веществом. Молекулы воды активно участвуют в поверхностном взаимодействии молекул белка друг с другом и с другими веществами. Термообработка повышает внутреннюю энергию костной ткани, в том числе энергию молекул воды. Последние освобождаются от внешних связей и выходят в свободное состояние. Общая активность воды увеличивается, адгезионная прочность резко снижается. Дальнейшая термообработка, как видно из графиков, практически восстанавливает исходную прочность костной ткани. Но она меняет свой состав и структуру. Белки подверглись денатурации, молекулы воды, находившиеся в связанном с белками состоянии, освободились и вышли в раствор, влага внутри ткани сохранилась только в химически и частично физико-химически связанном виде. Возрастание прочности в таком состоянии костной ткани можно объяснить формированием более компактной формы молекул белка, созданием непосредственного межмолекулярного контакта между ними и другими веществами. В таком состоянии кость становится более хрупкой, так как практически исчезает микроподвижность между минеральными пластинками, межклеточное вещество становится «более жёстким».
В результате проведенных исследований и их анализа установлено, что прочность обезвоженного растительного сырья определяется адгезионным поверхностным взаимодействием его макроструктур и «кирпичиков» (частично обезвоженных растительных клеток). Поверхностные белки клеточных мембран взаимодействуют с белками соседней мембраны как непосредственно, так и с участием молекул воды и веществом межклетников, содержащим белки либо пектины. Объекты переработки растительного происхождения чаще всего в нативном состоянии представляют собой коагуляционные дисперсные системы, влага в которых находится во всех формах связи с макромолекулами биополимеров. Высушивание создает условия для перехода структур из коагуляционных, когда поверхностное взаимодействие микроструктур, в частности – растительных клеток, осуществляется адгезионно (дисперсная фаза – дисперсионная среда), в конденсационно-кристаллизационные. Точку перехода растительной ткани из состояния коагуляционной структуры в конденсационно-кристаллизационную можно считать оптимальной для разрушения материалов растительного происхождения.
Библиографическая ссылка
Руднев С.Д., Киреев В.В., Попов Д.М. ОСОБЕННОСТИ ДЕФОРМАЦИОННОГО ПОВЕДЕНИЯ БИОЛОГИЧЕСКИХ ТКАНЕЙ // Современные наукоемкие технологии. 2015. № 3. С. 81-86;URL: https://top-technologies.ru/ru/article/view?id=34945 (дата обращения: 03.07.2026).