



Таблица 1. Некоторые показатели развития личинок и размеров сеголеток озерной лягушки при различном монохроматическом освещении
|
Цвет освещения |
Продолжительность (x+sx), сутки |
Отход метаморфизирующих особей, % |
Размеры сеголеток (x+sx) |
Индексы (x+sx), % от массы тела |
|||
|
личиночного развития |
метаморфоза |
длина, мм |
масса, мг |
печени |
сердца |
||
|
Контроль |
69.7+0.7 |
4.0+0.2 |
0 |
18.3 + 0.4 |
598 + 27 |
2.67 + 0.04 |
0.33 + 0.02 |
|
Красный |
67.5+0.9 |
4.3+0.4 |
2.5 |
17.9 + 0.3 |
586 + 36 |
2.57 + 0.06 |
0.33 + 0.02 |
|
Желтый |
65.3+0.6* |
3.6+0.3 |
0 |
19.2 + 0.2 |
726 + 26* |
2.74 + 0.03 |
0.46 + 0.01*** |
|
Зеленый |
65.5+0.6* |
3.5+0.2 |
0 |
19.6 + 0.3* |
762 + 28* |
2.69 + 0.07 |
0.46+0.02*** |
|
Голубой |
64.2+ 0.7** |
3.5+0.1* |
0 |
20.3 + 0.4** |
832 + 20** |
2.78 + 0.03* |
0.49 + 0.01*** |
|
Синий |
64.8+ 0.5** |
3.8+0.2 |
0 |
19.4 + 0.2 |
631 + 30 |
2.62 + 0.05 |
0.45 + 0.01** |
* Различия достоверны при P<0.05;
** Различия достоверны при P<0.01;
*** Различия достоверны при P<0.001.
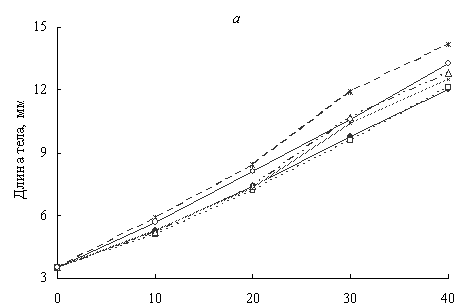
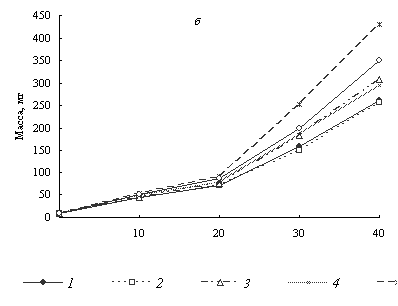
Рисунок 1. Динамика длины (а) и массы (б) тела личинок озерной лягушки в различных монохроматических режимах: 1 - контроль, 2 - красный, 3 - желтый, 4 - зеленый, 5 - голубой, 6 - синий.
По осям абсцисс - дни от вылупления.
Икра озерной лягушки Rana ridibunda получена из естественного водоема. До начала активного питания личинок, также как и икру, содержали в аквариуме объемом 100 л с принудительной аэрацией при непрерывном освещении люминесцентными лампами белого света (освещенность 100 лк). Эксперимент начали на личинках из одной кладки, которые находились на стадии 24 [11]. Опыты проводили в аквариумах емкостью 30 л со столбом воды 20 см. Плотность посадки 2 ос./л. Температура в опытах составляла 210С, содержание кислорода в воде 7.0-7.5 мг/л. Опытные аквариумы освещали различными зонами спектра. Характеристики всех светофильтров приведены ранее [8, 9]. Личинок кормили измельченными и ошпаренными листьями крапивы. Длину тела, массу и стадии развития [11] особей регистрировали через 10 дней. Общее физиологическое состояние сеголеток оценивали по морфофизиологическим показателям (индексам сердца и печени) в контроле и опыте [1]. Статистическая обработка цифрового материала проводилась с использованием t-критерия Стьюдента [3].
На рисунке 1 видно, что рост головастиков при красном освещении практически не отличался от контроля. При желтом и зеленом освещении длина тела особей на 40-й день выращивания превышала контрольную группу соответственно на 6.7 и 5.0% (различия недостоверны), масса - на 18.0 и 13.1% (P < 0.05). Более заметный эффект наблюдался при синем свете. По нашим данным благоприятным для роста головастиков озерной лягушки являлся голубой свет, при котором длина особей увеличилась на 18.3, а масса - на 64.7% по сравнению с контролем (P < 0.01). Коэффициент вариации длины и массы личинок снижался в аквариумах с освещением голубыми и синими лучами спектра. В остальных вариантах изменений не наблюдалось (табл.).
Как и ранее в опытах с травяной лягушкой [9], стадии развития головастиков мало отличались от контрольного варианта. Выживаемость личинок до наступления метаморфического климакса также была одинаковой и составила 100%. Длительность личиночного периода снижалась при любом монохроматическом освещении (табл.). Но если при красном свете этот показатель уменьшался относительно контроля недостоверно, то при голубом и синем освещении был достоверно ниже, соответственно на 7.9 и 7.0% (P < 0.01). Менее значимая разница по длительности развития зафиксирована в условиях желтого и зеленого освещения. В отличие от длительности развития продолжительность непосредственно метаморфоза (метаморфического климакса) не имела статистически достоверных различий. Исключение составлял голубой свет, при котором эта величина снижалась на 12.5% по сравнению с контролем (P < 0.05).
Во время метаморфоза в одном варианте с красным освещением наблюдалась гибель особей, в других аквариумах подобного не зафиксировано (табл.). Размеры сеголеток, выращенных в условиях голубого и зеленого освещения, были достоверно (P < 0.01, P < 0.05) выше таковых контрольных особей: по длине на 10.9 и 7.1%, по массе соответственно на 39.1 и 27.4%. Интересно, что линейные размеры сеголеток в других вариантах достоверно не отличались от размеров контрольных особей, а весовые показатели при желтом и зеленом освещении были достоверно выше (P < 0.05). Изучение морфофизиологических индексов показало, что индекс печени достоверно повысился у сеголеток, прошедших метаморфоз только в одном варианте - при голубом освещении. Напротив, индекс сердца при любом цвете освещения, кроме красного, достоверно увеличивался.
В умеренных зонах озерная лягушка откладывает икру обычно в тех местах, где температура для кладки наиболее благоприятна. После икрометания комки икры набухают и опускаются на дно между водными растениями. По нашим наблюдениям обычно глубина в подобных местах составляет 50-80 см. Вылупившиеся личинки расселяются по всему водоему, но всегда придерживаются зарослей подводной растительности, в которых они активно питаются. В таких условиях отдельные монохроматические зоны еще не поглощаются водной толщей и поэтому должны оказывать предположительно равнозначное действие на головастиков озерной лягушки.
Аналогично другим земноводным личинки озерной лягушки обладают цветовым зрением благодаря хорошо выраженным латеральным глазам и теменному органу [4]. При наименее комфортном освещении, красном свете, рост и развитие личинок, размеры сеголеток были хуже, чем в других монохроматических режимах. Мы уже высказывали гипотезы о механизмах влияния красной зоны спектра на личинок шпорцевой лягушки [8]. В полной мере они могут быть приложимы и к другим видам (травяной и озерной лягушкам), поскольку полученные результаты одинаковы для трех видов. Особо стоит выделить роль мелатонина, который, предположительно, выделяется в большом количестве при красном освещении. Этот гормон значительно подавляет пролиферацию клеток щитовидной железы, секрецию тироксина и, как следствие, понижает скорость развития [12, 17].
Подобно другим видам личинки Rana ridibunda лучше росли и быстрее развивались при зелено-синем освещении. При этом благоприятнее действовала голубая зона спектра. Наше предположение [8] об усилении синтеза соматотропина при освещении голубыми лучами подтверждается увеличением массы печени сеголеток, т.к. известна [7] роль этого гормона в усилении синтеза различных веществ в данном органе. К этой гипотезе можно добавить и предположение об антистрессовой направленности голубого света. Например, на нильской тиляпии [15] было показано, что при голубом освещении после стрессового воздействия содержание кортизола в плазме становится несколько меньше и не увеличивается, как при обычном освещении, т.е. в данном случае голубая зона спектра выступает в качестве стресс снимающего агента.
В отличие от голубого освещения, при синем, несмотря на достоверное увеличение ростовых показателей, размеры сеголеток были значительно меньше. Ранее [9] мы указывали, что при синем освещении личинки травяной лягушки развиваются значительно быстрее, однако во время метаморфоза при возрастании энергетических трат масса вышедших сеголеток снижается. Вероятной причиной в данном случае может являться усиление синтеза тироксина, резко увеличивающего энергетический обмен проходящих метаморфоз личинок. Например, он повышает активность дыхательных ферментов, синтез РНК и АТФ, а также стимулирует образование некоторых белков, характерных для взрослого организма (гемоглобин, сывороточный альбумин), которые затем поступают в кровяное русло [7, 14]. Из таблицы видно, что при синем освещении на фоне быстрого развития озерной лягушки масса печени ниже, чем в контроле, а общая масса сеголеток несколько выше (различия недостоверны). В то же время при сопоставлении полученных результатов с ранними нашими исследованиями прослеживается определенная зависимость. Синее освещение ускоряет развитие, но мало изменяет или снижает размеры сеголеток, что может быть связано с усилением синтеза тироксина.
Работа выполнена при поддержке гранта Президента РФ (МК-1066.2003.04).
СПИСОК ЛИТЕРАТУРЫ
- Ищенко В.Г. //Руководство по изучению земноводных и пресмыкающихся. Киев: Наукова думка, 1989. С. 92.
- Константинов А.С. Общая гидробиология. М.: Высш. школа, 1979. 480 с.
- Лакин Г.Ф. Биометрия. М.: Высш. школа, 1990. 293 с.
- Мантейфель Ю.Б. Зрительная система и поведение бесхвостых земноводных. М.: Наука, 1977. 266 с.
- Раденко В.Н., Алимов И.А. //Вопр. ихтиологии. 1991. Т. 31. Вып. 4. С. 655.
- Раденко В.Н., Терентьев П.В. //Биология сиговых рыб. М.: Наука, 1988. С. 216.
- Розен В.Б. Основы эндокринологии. М.: Изд-во МГУ, 1994. 384 с.
- Ручин А.Б. //Зоол. журнал. 2002. Т. 81. № 6. С. 752.
- Ручин А.Б. //Изв. РАН. Сер. биол. 2003. № 5. С. 582.
- Ручин А.Б., Вечканов В.С., Кузнецов В.А. //Вопр. ихтиологии. 2002. Т. 42. № 2. С. 236.
- Терентьев П.В. Лягушка. М.: Сов. наука, 1950. 345 с.
- Edwards M.L., Pivorun E.B. //Gen. and Comp. Endocrinol. 1991. V. 81. № 1. P. 28.
- Jinho Ch., Shuhei N. //Prelim. Rept Hakuho Maru Cruise. 1995. Kh-94-2. C. 57.
- Tata J.R. //Biochimie. 1999. V. 81. № 4. P. 359.
- Volpato G.L., Barreto R.E. //Brazil. J. Med. Biol. Res. 2001. V. 34. P. 1041.
- Wang F., Dong Sh., Huang G., Wu L., Tian X., Ma Sh. //Aquaculture. 2003. V. 228. № 1-4. P. 351.
- Wright M.L., Cuthbert K.L., Donohue M.J., Solano S.D., Proctor K.L. //J. Exp. Zool. 2000. V. 286. № 6. P. 625.
Библиографическая ссылка
Ручин А.Б., Лобачев Е.А. ВЛИЯНИЕ МОНОХРОМАТИЧЕСКОГО СВЕТА НА РОСТ И РАЗВИТИЕ ЛИЧИНОК ОЗЕРНОЙ ЛЯГУШКИ, RANA RIDIBUNDA // Современные наукоемкие технологии. 2005. № 8. С. 13-16;URL: https://top-technologies.ru/ru/article/view?id=23442 (дата обращения: 30.06.2026).