



Зрительная система, как и любой исследуемый объект, подвергается изучению в четырех аспектах, касающихся структуры, происходящих процессов, изменения состояний и принципов управления. Применительно к зрительной системе эти аспекты иллюстрируются на рис. 1. Сведения о структуре системы, ее состояниях и специфических процессах, к которым относят рецепцию, анализ и регистрацию зрительной информации, а также – опознание образов и элементы мышления, лежат в основе знания об организации зрительной функции. Однако нельзя забывать о самом важном аспекте – выяснении принципов функциональной организации и управления зрительной системой в рамках поведения, цель которого выживание и удовлетворение потребностей организма. Неспецифические процессы, в отличие от специфических, обусловленных конкретным внешним воздействием, выражают вовлечение нервных структур в целостную мозговую деятельность, определяемую оценкой поведенческой ситуации и прогнозом ее развития. Предполагается, что функциями управления в соответствии с потребностями и мотивациями наделен особый мозговой аппарат, осуществляющий, в частности, эмоциогенез, регуляцию состояний в континууме сон-бодрствование, формирование неизбирательного и избирательного внимания. Понятие «неспецифические системы» [1] применяется для определения этого аппарата с учетом множественности мозговых образований, входящих в его состав. Термин «неспецифические образования мозга» определяет распределенные в пространстве мозга нервные структуры и нейронные комплексы, которые выполняют функцию управления специфическими процессами.
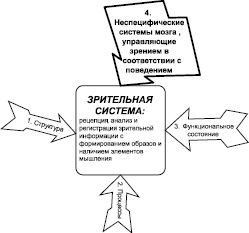
Рис. 1. Аспекты изучения зрительной системы
Сенсорная информация, поступающая в центральные отделы зрительной системы, представлена потоками дискретных посылок, отличающихся по сигнальной значимости. Начиная с классических исследований [8] сохраняется представление о том, что неспецифический контроль над поступающей информацией осуществляется стволовой ретикулярной формацией (formatio reticularis, RF), ответственной за регуляцию функционального состояния специфических отделов мозга. Однако при этом остается открытым вопрос, как производится управление обработкой множества отдельных дискретных частиц информации, составляющих целостные образы и используемых для принятия поведенческих решений.
Цель данной работы состояла в получении сведений о функциональной значимости неспецифических образований кортикальных отделов зрительной системы и в объяснении этих сведений на основе гипотезы о гетерогенности нейронных модулей, обеспечивающих анализ и регистрацию сенсорной информации. Для достижения цели решали ряд задач. Во-первых, выясняли минимальную интенсивность электрического раздражения пунктов коры больших полушарий и RF среднего мозга для возникновения реакции активации мозга у интактных животных и в условиях снижения влияния RF. Во-вторых, сравнивали пороги реагирования на вызванное светом возбуждение, поступившее по основному зрительному тракту при отключении зрительных путей в RF, с порогами реагирования в условиях разрушения основного зрительного тракта при сохранении коллатеральных нервных каналов, обеспечивающих RF зрительным возбуждением. В-третьих, оценивали функциональную значимость неспецифических кортикальных образований, сравнивая обучение дифференцировать зрительные сигналы интактных животных и тех, у которых кора больших полушарий лишалась ретикулярных влияний, обусловленных зрительным возбуждением.
Материалы и методы исследования
Принципиальная особенность всех экспериментов состояла в том, что в них использовали бодрствующих животных (кошки, самцы весом 3,5–4,0 кг), что позволяло следить за их поведением и изменением уровня активации мозга. Наблюдали за двумя группами животных с разрушениями структур мозга (иллюстрируются на рис. 2), а также за группой интактных животных.
Для получения первого препарата, обозначенного стрелкой 1 на схеме на рис. 2, у 12 животных посредством двусторонней полной электрокоагуляции наружных коленчатых тел (corpus geniculatum laterale, CGL) разрушали основной зрительный путь в кору, не повреждая коллатеральные нервные волокна в RF. Второй препарат (отмечен, стрелкой 2, рис. 2) создавали посредством электрокоагуляции у 28 животных ручек верхних бугров четверохолмия (colliculus superior, CS). Тем самым разрушали коллатеральные зрительные пути в RF, не затрагивая основной канал зрительного сенсорного возбуждения в кору. Животным обеих групп с повреждениями мозга, а также – интактным, в ходе операции, проводившейся с соблюдением биоэтических норм в стерильных условиях под наркозом (нембутал 25 мг на кг веса, внутрибрюшинно), вживляли в мозг (в RF, дорзальный отдел CGL, в 17, 18 и 19 поля зрительной коры, а также в лобную и височную кору) электроды, изготовленные из нихромовой проволоки диаметром 0,1 мм в стеклянной изоляции. В начальной стадии экспериментов регистрировали пороговую интенсивность светового (засветка матового диска перед глазами животного 3 с, 30–250 лк) и электрического (биполярные импульсы 1 мс, 200 Гц) раздражения для вызова реакции активации ЭЭГ. Затем, подкрепляя световой сигнал 30 лк кусочком мяса, сравнивали скорость выработки условного рефлекса у животных с разрушением ручек CS и у интактных животных.
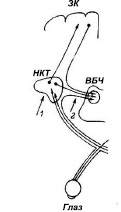
Рис. 2. Схема препаратов мозга с деструкцией зрительных путей у животных двух экспериментальных групп:1 – блокада основного зрительного пути в зрительные отделы неокортекса (ЗК) посредством двусторонней электрокоагуляции наружных коленчатых тел (НКТ);2 – разрушение зрительных коллатералей в ретикулярную формацию посредством двусторонней электрокоагуляции ручек верхних бугров четверохолмия (ВБЧ)
Результаты исследования и их обсуждение
Решая первую экспериментальную задачу, сопоставляли пороги электрического раздражения пунктов мозга (см. схему на рис. 3) у животных без повреждений мозга и у тех, которые подверглись операции полного двустороннего разрушения ручек CS, обеспечивающих поступление зрительного возбуждения в RF.
Локализация на схеме рис. 3 больших кружков иллюстрирует присутствие в коре больших полушарий пунктов с низким порогом, не превышавшим 120 мкА. Такие пороги соответствовали результатам измерения порогов раздражения RF, находившихся в пределах 40–120 мкА. Совокупные данные по исследованным точкам приведены в таблице.
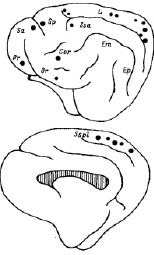
Рис. 3. Контурная схема левого полушария головного мозга кошки, вид с двух сторон (заштриховано мозолистое тело). Кружки разного диаметра отмечают локализацию кортикальных пунктов, в которых измеряли пороги электрического раздражения для вызова активации мозга. Схема соответствует нумерации пунктов табл. 1: 1 - прореальная извилина (Pr); 2, 3 - передняя и задняя сигмовидные (Sa, Sp); 4 - передняя супрасильвиева (Sca); 5,6 - оральный и каудальный отделы латеральной извилины (L); 7 - коронарная (Cor); 8 - орбитальная (Or); 9 - супрасплениальная извилина (Scpl). Диаметр кружка обратно пропорционален величине порога: наибольшие кружки для низких порогов 40–120 мкА, которые соответствовали RF, средние – для порогов 120–220 мкА и наименьшие – для порогов свыше 220 мкА
Обращает на себя внимание характер изменения порогов после операции разрушения зрительных коллатералей в RF. Ожидалось повышение порогов, поскольку понизилось зрительное возбуждение RF и, соответственно, уменьшилось активирующее влияние RF на кору. Однако, наоборот, пороги понизились. На рис. 3 не разделяются результаты наблюдений на интактных и оперированных животных, потому что пороги и тех и других оставались в одних и тех же условно выделенных пределах.
При решении второй исследовательской задачи в экспериментах на кошках, у которых произвели разрушение основного зрительного пути (вариант 1 на схеме рис. 2) возникновение отчетливой десинхронизации ЭЭГ происходило при пороговом световом раздражении 250 лк, что на 2 порядка выше, чем у интактных животных (2 лк). Контролем функционального состояния животного (того, что именно дефицит зрительного возбуждения явился причиной изменения порогов) служил нормативный уровень пороговой активации на слуховое раздражение (тон 1000 Гц, 10 дБ). Вместе с тем, отметили, что животные с разрушением зрительных коллатералей в RF (вариант 2 на рис. 2), и интактные животные неизменно отвечали реакцией активации ЭЭГ на пороговые световые вспышки интенсивностью 2 лк. Тем самым было показано, что неспецифические образования, находящиеся в коре, достаточно эффективны для поддержания активации в условиях падения влияния из RF. Отмеченное после двусторонней электрокоагуляции CGL снижение реактивности указывало на то, что основной зрительный путь (поражение которого обусловило падение реактивности) и те неспецифические образования, которые снабжались возбуждением по этому пути, обладают гораздо большим активационным потенциалом, чем RF получившая возбуждение по коллатеральным зрительным каналам.
Пороговые величины электрического раздражения пунктов коры для вызова активации мозга у интактных животных (А) и при сниженном уровне ретикулярных влияний (Б)
|
Номер раздражаемой точки (см. рис. 3) |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
|
А. Средняя величина порогов и доверительные интервалы (мкА) |
110 ± 8 |
120 ± 5 |
115 ± 9 |
198 ± 12 |
157 ± 8 |
115 ± 5 |
91 ± 10 |
156 ± 17 |
120 ± 4 |
|
Б. Средняя величина порогов и доверительные интервалы (мкА) |
96 ± 5 |
93 ± 9 |
90 ± 7 |
172 ± 11 |
140 ± 8 |
105 ± 3 |
89 ± 8 |
154 ± 14 |
110 ± 5 |
|
Отношение величины понижения порога к значению порога в норме (% %) |
13 % |
22 % |
22 % |
13 % |
11 % |
8 % |
2 % |
1 % |
8 % |
В наблюдениях, связанных с решением третьей экспериментальной задачи, не выявили нарушений в зрительной ориентации у животных, подвергшихся операции разрушения ручек CS. При этом отмечено достоверное ухудшение памяти у животных с полным разрушением ручек CS и замедление скорости обучения. Им требовалось 48 ± 7 тренировочных опытов для выработки пищедобывательной реакции на подкрепляемый световой сигнал, в то время как контрольные (интактные и с неполным разрушением) животные закрепляли рефлекс в памяти после 35 ± 3 повторений.
При регистрации ЭЭГ реакции была подтверждена ранее установленная закономерность [2], выраженность ответной реакции находилась в зависимости от биологической значимости сигнала. Важно отметить, что зависимость проявилась вопреки закономерности, определявшей характер ответа при действии безусловных стимулов. Имеется в виду, что положительный сигнал был самым слабым по физической интенсивности (30 лк), а отрицательные (250 и 125 лк) – более сильными.
Приведенные данные, продемонстрировавшие функциональную эффективность неспецифических образований коры больших полушарий и вовлеченность в работу памяти, вместе с литературными сведениями о влиянии активации мозга на анализ и регистрацию зрительной информации, в частности [6, 7], создали предпосылку для гипотезы о сопряжении специфических и неспецифических нейронов в структуре гетерогенного модуля, обеспечивающего анализ и регистрацию получаемой информации. На рис. 4 приводится схема, иллюстрирующая два варианта гипотез о соотношении регуляторов и регистраторов в работе памяти. Первый вариант (1) предусматривает пространственное обособление блока управления от блока регистрации. Второй вариант (2) предполагает совмещение и сопряжение в едином модуле специфических и неспецифических нейронов. Благодаря сопряженности обеспечивается детальное и надежное управление запоминанием, анализом и воспроизведением фрагментов информации. Кроме того, сопряженность объясняет работу механизма целесообразной фрагментации сенсорной информации [3, 4]. Имеется в виду, что создается запрос (определяется комплекс информации, которая требуется для принятия поведенческого решения), а затем происходит селекция биологически значимых сигналов и фрагментация потоков доступной информации в соответствии с весом, обусловленным запросом [5]. Можно полагать, что неспецифические нейроны, вынесенные в сенсорные и ассоциативные отделы мозга, находятся в подчинении по отношению к тем, которые находятся в лимбико-ретикулярных структурах, составляющих центральное звено неспецифических систем мозга, то есть, комплекса корковых и подкорковых образований, обеспечивающих регуляцию работы мозга и сомато-вегетативных систем организма в процессе поведения [1].
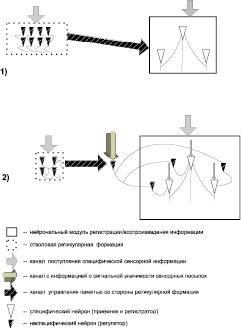
Рис. 4. Схемы двух вариантов управления памятью:первый (1) – за счет взаимодействия структурно обособленного регуляторного блока со специфическим регистрирующим блоком; второй (2)) – при пространственном совмещении специфических и неспецифических нейронов в модуле памяти
Чем выше вес информации (то есть, чем она значимее для поведения), тем выше качество, размеры и количество ячеек, отводимых для хранения и тем прочнее память – и наоборот, чем ниже вес, тем менее стабильна ячейка. По всей видимости, изменение качества и количества ячеек, занимаемых информацией, происходит под воздействием регулятора, пространственно совмещенного со специфическими нейронами. Особого рассмотрения заслуживает память, служащая буфером информации с таким малым весом, который не предполагает прочной регистрации. Имеется в виду эпизодическая и промежуточная память. Наличие такого буфера перестает выглядеть избыточным, если учесть, что содержащаяся в нем информация, поддающаяся извлечению в стрессовой ситуации, повышает вероятность выживания в условиях кризисных обстоятельств. Предполагается, что использование модельных представлений, в сочетании с изучением алгоритмов манипуляции весовыми параметрами доступной информации будет содействовать решению проблемы возмещения дефицита достоверной информации в процессе формирования целостных образов и поможет понять природу иллюзий.
Заключение
В результате проведенного исследования, показавшего сопряжение специфических и неспецифических нейронов в кортикальных нервных сетях и функциональность отмеченного сопряжения при формировании памяти в ходе выработки условного рефлекса, предложена принципиально новая модель гетерогенного модуля памяти. Модель, учитывающая дискретность воспринимаемой и регистрируемой информации, а также распределение памяти по категориям (буферной, краткосрочной и долговременной), отражает работу механизма, управляющего регистрацией фрагментов информации в ячейках памяти. Согласно представленной модели, переход памяти из одной категории в другую обусловлен весовыми параметрами информации и обеспечивается изменением состояния и структуры ячеек, в которых хранятся фрагменты информации. Изменение состояния происходит под контролем со стороны неспецифических нейронов, совмещенных со специфическими в едином функциональном модуле. Имеется в виду, что неспецифические нейроны гетерогенного модуля выполняют двойную функцию, с одной стороны, контролируют работу специфических нейронов, а с другой стороны, включены в целостную деятельность неспецифических систем мозга, ответственных за регуляцию и консолидацию мозговых процессов.